Using Decision Analysis to Determine the Feasibility of a Conservation Translocation
Abstract
Conservation translocations, intentional movements of species to protect against extinction, have become widespread in recent decades and are projected to increase further as biodiversity loss continues worldwide. The literature abounds with analyses to inform translocations and assess whether they are successful, but the fundamental question of whether they should be initiated at all is rarely addressed formally. We used decision analysis to assess northern leopard frog reintroduction in northern Idaho, with success defined as a population that persists for at least 50 years. The Idaho Department of Fish and Game was the decision maker (i.e., the agency that will use this assessment to inform their decisions). Stakeholders from government, indigenous groups, academia, land management agencies, and conservation organizations also participated. We built an age-structured population model to predict how management alternatives would affect probability of success. In the model, we explicitly represented epistemic uncertainty around a success criterion (probability of persistence) characterized by aleatory uncertainty. For the leading alternative, the mean probability of persistence was 40%. The distribution of the modelling results was bimodal, with most parameter combinations resulting in either very low (<5%) or relatively high (>95%) probabilities of success. Along with other considerations, including cost, the Idaho Department of Fish and Game will use this assessment to inform a decision regarding reintroduction of northern leopard frogs. Conservation translocations may benefit greatly from more widespread use of decision analysis to counter the complexity and uncertainty inherent in these decisions.
History: This paper has been accepted for the Decision Analysis Special Issue on Decision Analysis to Advance Environmental Sustainability.
Funding: This work was supported by the Wilder Institute/Calgary Zoo, the U.S. Fish and Wildlife Service [Grant F18AS00095], the NSF Idaho EPSCoR Program and the National Science Foundation [Grant OIA-1757324], and the Hunt Family Foundation.
Supplemental Material: The online appendix is available at https://doi.org/10.1287/deca.2023.0472.
Introduction
Conservation translocations, including reintroduction and reinforcement, are increasingly being used as management tools to mitigate species declines (Seddon et al. 2007, Germano and Bishop 2009, Harding et al. 2016). Reintroduction involves the intentional movement and release of wild or captive-bred individuals back into their indigenous range following extirpation (i.e., local extinction), whereas reinforcement involves the same actions but into an extant (i.e., still in existence) population of conspecifics (IUCN/SSC 2013). Reintroduction and/or reinforcement have been used in conservation programs in North America for at least 279 animal species and, more specifically, at least 52 species of reptiles and amphibians, with the number of publications related to conservation translocations steadily rising since the 1970s (Brichieri‐Colombi and Moehrenschlager 2016). Although rare in practice to date, translocation is commonly proposed as a climate-change adaptation tool (Butt et al. 2021) and will likely continue to increase in light of climate change and ongoing global biodiversity loss (Swan et al. 2018).
Conservation translocation programs often involve difficult decisions regarding how and when to act in response to threats impacting at-risk species and ecosystems. Translocation programs typically require considerable resources (e.g., staffing, captive breeding program development and maintenance, postrelease monitoring; Canessa et al. 2014, Kissel et al. 2017, Hilbers et al. 2019; for a discussion of costs specific to this assessment, see Randall et al. 2021) and invariably result in those resources not being allocated to other conservation actions or species (Martin et al. 2018). Therefore, a proactive evaluation of likelihood of success prior to investing in a conservation translocation is a critical, but generally lacking, step in deciding whether conservation resources should be devoted to translocation of a particular species. More widespread use of decision analysis to counter the complexity and uncertainty inherent to these decisions would help maximize the effectiveness of conservation resources in conservation decisions (Martin et al. 2018).
Decision analysis is a systematic process for decision making that involves deconstructing a decision into the key components (Howard 1966, Howard and Abbas 2015; see Hemming et al. 2022 for a conservation example). Carrying out the steps of a decision analysis can encourage a more precise thought process regarding the nature of the decision, the fundamental objectives one seeks to achieve, the alternative actions for addressing the problem, potential outcomes of each alternative, and the relative value of each outcome in terms of the fundamental objectives (Hemming et al. 2022). Although there are a plethora of examples in the literature using decision analyses to inform translocation activities and assess whether conservation actions are successful (Converse et al. 2013, Canessa et al. 2014, Kissel et al. 2017, Seaborn and Goldberg 2020, Soorae 2021), the fundamental question of whether they should be initiated at all is rarely assessed as a formal decision.
The State Wildlife Action Plan for Idaho recommended conducting a feasibility assessment for northern leopard frog (Lithobates pipiens) recovery options in northern Idaho (Idaho Department of Fish and Game 2017). Although northern leopard frogs are one of the most widely distributed amphibian species in North America, they have experienced significant declines in the Pacific Northwest in recent decades (British Columbia: COSEWIC 2009; Washington: Germaine and Hays 2009; Idaho: Lucid et al. 2016; Oregon: Rorabaugh 2005). The species is currently extirpated from its historical range in northern Idaho (Lucid et al. 2020) and is classified as imperiled in the state (NatureServe 2022). The nearest remaining population of northern leopard frogs is located at the Creston Valley Wildlife Management Area, 22 kilometers north of the U.S.–Canadian border; this comprises the only extant population in British Columbia and is federally listed as endangered in Canada (COSEWIC 2009). Northern leopard frogs also occur in southern Idaho, but these populations are genetically distinct from the British Columbia population (Hoffman and Blouin 2004).
We undertook a decision-analytic process to assess the feasibility of northern leopard frog reintroduction in northern Idaho (Randall et al. 2021). The assessment was initiated in 2019 but the bulk of the work occurred primarily in winter 2020/spring 2021 over a period of six months. Stakeholders, titleholders, and subject matter experts from government, indigenous groups, academia, conservation organizations, and land management agencies were involved in the problem framing, specification of objectives, and development of alternatives. We built an age-structured population model to predict how the different management alternatives would affect the probability of reintroduction success. Age-structured population models are a common approach to population modeling; these models are used to forecast the abundance of each age class while allowing for age-specific vital rates (e.g., survival and reproduction; Tuljapurkar and Caswell 1997, Gerber and González-Suárez 2010). The results of the analysis will inform the decision maker and stakeholders of the likelihood of a successful reintroduction and will provide a modeling framework that may support future analyses of this decision.
Methods
We used a decision-analytic framing for our feasibility assessment (Canessa et al. 2018). Specifically, we followed the PrOACT approach (i.e., problem framing, objectives, alternatives, consequences, and tradeoffs; Hammond et al. 1999). We carried out our assessment over a six-month period with the participation of a group of 14 people, including representatives of the decision-making agency, stakeholders, experts, and one decision analyst (for full details, see Randall et al. 2021). Some of the experts and representatives of the decision-making agency, as well as the decision analyst, are coauthors of this paper.
Problem Framing and Objectives
The problem framing and objectives steps of the feasibility assessment were conducted primarily during six one-hour virtual issue-raising sessions (Tani and Parnell 2013b, Howard and Abbas 2015). These sessions were facilitated by the decision analyst. In these sessions, participants first discussed their questions and concerns regarding northern leopard frog reintroduction in northern Idaho in an open brainstorming format resulting in a list of relevant issues. Once the issues were identified, the facilitator guided the discussion to sort the issues into four categories: (1) decisions, (2) objectives, (3) uncertainties, and (4) other (process issues or facts).
The decisions from the issue-raising sessions were further sorted into a decision hierarchy (Tani and Parnell 2013b, Howard and Abbas 2015) using three categories: already decided/assumptions, focal decisions, and issues to be decided later (Figure 1). Already decided/assumptions are decisions that have already been made or are being assumed for the purposes of this assessment. Focal decisions are those to consider further in the assessment. Issues to be decided later are important but can be addressed once the main strategy is chosen. The decision hierarchy clarified assumptions and identified focal decisions that were subsequently the focus of our analysis. The decision hierarchy was also useful for getting alignment on the scope of the assessment. For example, the study area was initially much larger. Discussions during the issue-raising sessions led to constraining the study area to the Boundary Smith Creek Wildlife Management Area (BSCWMA; Figure 2). This decision was then categorized as already decided. The BSCWMA covers 8.4 km2 (2,072 acres) in the Kootenai River valley directly adjacent to the U.S.–Canadian border (Trese 2014). The numbered cells in the BSCWMA, as shown in Figure 2, refer to wetland names that are later referenced as potential reintroduction locations. The Creston Valley Wildlife Management Area (CVWMA) in British Columbia (Figure 2) represents the closest northern leopard frog population to the area of interest.
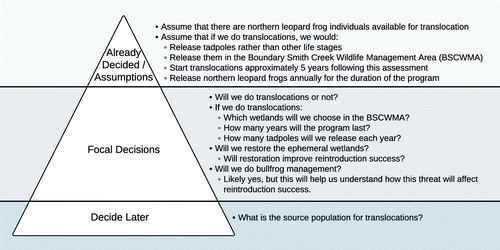
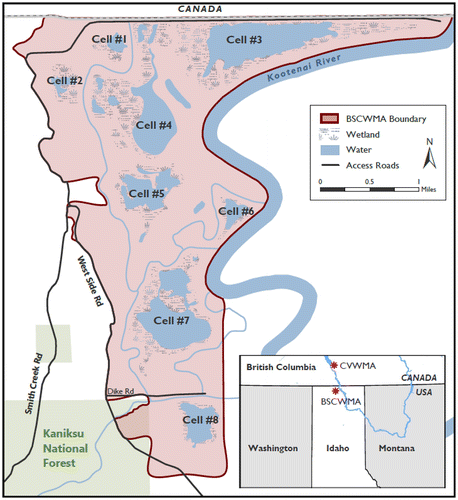
Source. Adapted from Idaho Department of Fish and Game.
Note. CVWMA, Creston Valley Wildlife Management Area.
We initially identified two fundamental objectives: (1) recovery of northern leopard frogs in northern Idaho and (2) minimizing cost. We defined recovery of northern leopard frogs as the establishment of a population that persists for at least 50 years, measured as a probability of persistence that we sought to maximize. Probability of persistence is a metric that is often used in conservation biology to characterize objectives around preventing extinction because it captures the aleatory uncertainty inherent in natural systems (Gerber and González-Suárez 2010). We did not define an acceptable threshold for probability of persistence in advance for two reasons: (1) we wanted the assessment to be useful into the future (a decision around the reintroduction would not happen immediately) and therefore wanted it to withstand changes in what is acceptable for the decision maker, and (2) we were concerned that a focus on a single threshold could distract from useful insights. Maximizing the number of frogs was also discussed as a possible measurable criterion but was ultimately not used since it did not capture the intent of the recovery objective. For example, if there are two alternatives and both result in a healthy population of northern leopard frogs then the two alternatives would be equivalent at achieving the fundamental objective of northern leopard frog recovery, even if one had more frogs.
Although we recognized minimizing cost as a fundamental objective, here we focused solely on the recovery objective within the context of management in the BSCWMA. We agreed that costs could be quantified later if the recovery results looked promising. We decided not to include an assessment of cost in this initial analysis for two reasons: (1) a cost analysis would not be necessary if the results of the assessment for the recovery objective were not encouraging, and (2) our decide later decision around the source population meant that it would be difficult to estimate that aspect of the cost given that the source population could have major implications for cost. The decision around source population was categorized as decide later in the decision hierarchy (Figure 1) because there was considerable debate among the stakeholders regarding the pros and cons of various possibilities but ultimately it did not matter for the assessment of the recovery objective. By classifying it as decide later, we were able to move forward with the process. If cost is assessed in the future, the source population could become a focus on decision so that alternatives can be clearly specified and the corresponding costs quantified.
The ultimate decision makers were the Idaho Department of Fish and Game senior leadership and a citizens’ management committee. Neither was directly involved in this assessment. For the purposes of this feasibility assessment, we considered the decision maker to be the representative from the Idaho Department of Fish and Game who initiated the assessment and makes recommendations to the ultimate decision makers. One limitation of this approach is that directly involving the true decision makers in the process could increase the chance that they find the assessment useful (Gregory et al. 2012).
Ultimately, the framing exercise resulted in the following goal statement to summarize the purpose of the feasibility assessment: The Idaho Department of Fish and Game seeks to gain insight into the likelihood of success of reintroducing northern leopard frogs in the BSCWMA in northern Idaho to achieve local recovery of the species, given that the species is currently extirpated from the region.
Alternatives
The focal decisions identified in the decision hierarchy formed the columns of a strategy table (Tani and Parnell 2013a, Howard and Abbas 2015) that was used to guide brainstorming on alternatives over three one-hour virtual sessions. Four management alternatives were constructed from the options identified based on varying levels of effort for a northern leopard frog reintroduction program (Table 1).
|

Table 1. Alternative Management Strategies for Reintroduction of Northern Leopard Frogs into the BSCWMA in Northern Idaho
| Alternative name | Tadpole releases | Release location | Restore the ephemeral wetlands | Bullfrog management | Number of years of releases | Number of tadpoles released per year |
|---|---|---|---|---|---|---|
| Do nothinga | No | None | No | No | 0 | 0 |
| Go Big or Go Homeb | Yes | Cells 3, 4, and 7 | Yes | Yes | 10 | 18,000 |
| Middle of the Roadc | Yes | Cells 3, 4, and 7 | No | Yes | 5 | 9,000 |
| Minimum funding/low effortd | Yes | Cell 7 | No | Yes | 2 | 2,500 |
aStatus quo.
bIncreased effort.
cMost realistic based on past efforts reintroducing northern leopard frogs in British Columbia.
dUsed to assess whether the low effort could lead to success.
The Middle of the Road alternative provides an example of what has been done in previous northern leopard frog reintroductions in British Columbia in terms of number of individuals and years of effort, whereas Go Big or Go Home represents twice this amount of effort along with restoration of ephemeral wetlands. Release locations (wetland cells) were selected based on prior knowledge related to suitability for northern leopard frogs; both the Middle of the Road and the Go Big or Go Home alternatives include release in three wetlands. The Minimum Funding/Low Effort alternative was included to assess whether a smaller number of individuals released and shorter period of reintroduction activities could still lead to success; given that this alternative included only 2500 tadpoles being released per year, it would be unnecessary to release in more than one wetland due to the low density of tadpoles that would be released to the wetland. Bullfrog (Lithobates catesbeianus; an invasive species that depredates and competes with local frogs) management was deemed to be necessary in all cases of reintroduction.
Consequences
We built an age-structured population model to simulate the demography of a reintroduced northern leopard frog population in the BSCWMA under each alternative. Population models such as this are widely accepted as useful for providing insights, particularly for relative comparisons of management alternatives, but caution is recommended when interpreting the absolute values of the results (Morris and Doak 2002, Gerber and González-Suárez 2010). Here we describe the model at a high level, but for additional modeling details refer to Randall et al. (2021).
The model structure incorporated the management objective, focal decisions, and uncertainties identified during the issue-raising sessions. An influence diagram (Howard and Matheson 2005) summarizing the overall model structure is provided in Figure 3. The age structure incorporated in the model is provided in Figure 4. The uncertainties in the model (single ovals in Figure 3) were parameterized with different probability distributions for each of the age classes shown in Figure 4, where relevant. We only represented females in the model because sex ratios for northern leopard frogs tend to be approximately 1:1 (Merrell 1968, Hine et al. 1981).
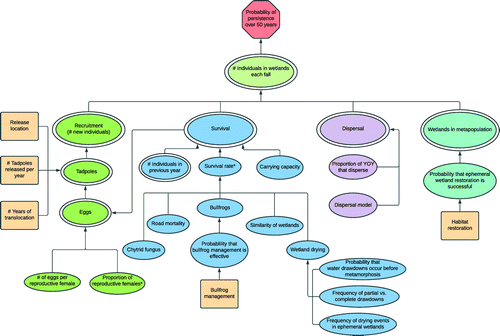
Notes. YOY, young of year. Octagon = objective, double oval = calculation, single oval = uncertainty, rectangle = decision. *Both an annual mean and standard deviation were elicited as uncertain parameters.
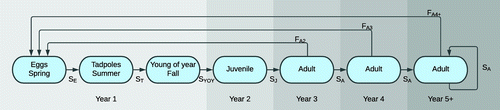
Note. S, survival rates; F, reproductive rates (fecundity); E, eggs; T, tadpoles; YOY, young of year; J, juvenile; A, adult.
Recruitment of northern leopard frogs was modelled as a function of both in situ reproduction and translocation of tadpoles into the BSCWMA. In situ reproduction was modeled as the proportion of reproductive females in each of the adult age classes and the average number of eggs per reproductive female, resulting in the number of eggs laid each year for each age class. Survival rates were applied to the eggs to determine the number of wild-born tadpoles. The number of translocated tadpoles (i.e., no. of tadpoles released) per year was added to the number of wild-born tadpoles in each wetland to reflect the total number of tadpoles in each wetland each year. The number of tadpoles released per year, the number of years of translocations, and the release location(s) varied by management alternative.
Survival of northern leopard frogs was modelled using survival rates and carrying capacity. Survival rates were modeled as a base survival rate for each life stage modified by threats. The base survival rate represents the survival rate for each life stage in the absence of threats such as chytrid fungus (Batrachochytrium dendrobatidis), road mortality, bullfrogs, and wetland drying. The impact of those threats on survival rates were modeled as a proportion reduction in the survival rate. Multiple threats were modelled multiplicatively, with the reduction in survival for each additional threat applied to the survival rate modified by all previously applied threats. For example, consider survival rate s and proportion reduction in survival due to bullfrogs and road mortality as x and y, respectively. Then the survival rates were modeled as s(1 − x)(1 − y). We set a lower bound of 1% on the annual survival rate for tadpoles based on feedback from our experts that, in their experience, generally at least some released tadpoles survive to become young of year. We modeled the similarity of wetlands as a correlation applied to the temporal variation of egg and tadpole survival rates for each wetland.
Carrying capacity was included as a simple ceiling on the population size of terrestrial age classes (i.e., young of year, juvenile, and adult life stages). If the population size of terrestrial age classes was greater than the carrying capacity, the population size of each terrestrial life stage was reduced proportionally to the population size of the life stage such that the total population size did not exceed the carrying capacity.
Dispersal was modeled as the proportion of young of year who disperse each year and the probability of moving each of the following three distance categories (given dispersal): (1) ≤1 km, (2) >1 km but ≤2 km, and (3) >2 km. Two possibilities for the probability of moving each of the three distance categories were incorporated: (1) Columbia spotted frog model, where individuals are more likely to go either short or long distances, and (2) More Go Short model, where individuals are more likely to go shorter distances. This structural uncertainty was represented probabilistically, with a probability assigned to each of the two dispersal models.
The wetlands in the metapopulation were modeled as the wetlands in the BSCWMA with suitable northern leopard frog habitat. Whether the ephemeral wetlands were considered suitable depended on the decision to do habitat restoration and the probability of successful restoration.
Expert elicitation was used to assign a probability distribution for each parameter in the model. The information came from either a single expert or pair of experts working together to produce a single estimate (i.e., the two people together were effectively a single expert). Experts used available information from the literature and their relevant practical experience to inform their judgements. The elicitation process for each parameter generally followed the five steps recommended by McNamee and Celona (2007): (1) motivating, (2) structuring, (3) conditioning, (4) encoding, and (5) verification. The motivating step was done by explaining the purpose and process to the experts. The structuring step was done in discussion with the experts to understand how the experts thought about the parameter, specify a clear definition, and uncover any assumptions. The conditioning step was done by having the experts explore extreme scenarios to counteract anchoring and availability biases. The encoding step is discussed further below in the following paragraphs depending on the type of variable. The verification step was done by providing the experts with graphs showing the resulting distribution for each of their estimates and giving them the opportunity to adjust. The facilitator walked the experts through a practice question in advance of the elicitation to help the experts become familiar with the elicitation format.
For parameters that could be represented as continuous variables (i.e., mean and standard deviation of proportion of reproductive females, mean number of eggs per female, mean and standard deviation of survival rates, proportion reduction in survival rate due to each threat, frequency of partial versus complete drawdowns, frequency of drying events in ephemeral wetlands, carrying capacity, similarity of wetlands, and proportion of young of year who disperse), experts identified best guess estimates and associated credible intervals to reflect epistemic uncertainty using an elicitation procedure where experts were asked for upper and lower estimates, a best guess, and a confidence estimate (Speirs-Bridge et al. 2010). The elicitation results were converted into quantiles of a continuous probability distribution by assuming that upper and lower estimates represented a central credible interval reflecting the confidence estimate (Hemming et al. 2020). The best guess was predefined for experts as the median (i.e., the 50th percentile). Experts were also asked to identify absolute bounds for the resulting distribution, if applicable. The bounds and quantiles for each variable were used to parameterize metalog distributions (Keelin 2016) that were used to represent epistemic uncertainty in the model.
Four independent parameters were elicited as discrete events with a single probability: (1) the probability that bullfrog management is effective, (2) the probability that management drawdowns will occur after metamorphosis, (3) the probability that frogs disperse using the Columbia spotted frog dispersal model (versus the More Go Short model), and (4) the probability that ephemeral wetland restoration is effective. High and low estimates were also elicited for these four parameters to help facilitate expert thinking but were not used for modelling purposes (Hanea et al. 2021).
The model was built as a discrete-time stochastic Markov process simulation model and followed the methods of McGowan et al. (2011) and White (2000) for incorporating both epistemic (parametric) uncertainty and aleatory (stochastic) uncertainty (both environmental and demographic). The model included three nested loops: (1) an iteration loop that used one parameter draw per iteration and calculated the probability of persistence as the proportion of runs where the frog population size was greater than zero, (2) a run loop that replicated the parameter draw at the iteration level for a specified number of times (restarting the simulation at time t = 0 for each run), and (3) an annual loop that simulated the dynamics of the system (i.e., survival and reproduction of frogs) over the time frame of the model (50 years). The model was implemented in the programming language R (R Core Team 2021).
We analyzed the model results for each alternative by plotting the probability of persistence, and uncertainty around it, both over time and in year 50. We used violin plots to display the distribution of results. We also used excess probability distributions (i.e., complementary cumulative distributions) to assess first-order stochastic dominance among the alternatives, which gives us insight into how important risk attitude is to the decision (Howard 1966, Howard and Abbas 2015, Canessa et al. 2016). An excess probability distribution shows us the probability that the value metric (i.e., probability of persistence) will exceed any specified number (Howard and Abbas 2015). An alternative with excess probabilities that are larger than that of another alternative for every value metric (and therefore the curves of the excess probability distribution do not cross) would be the rational choice regardless of the decision maker’s risk attitude. Additional information on stochastic dominance is provided in Appendix A.
We investigated model sensitivity to epistemic uncertainty for the leading alternative using an expected value tornado diagram (Johnson and Tani 2013, Howard and Abbas 2015). For each uncertainty, the parameter was held at either a low (≤10th percentile) or high (≥90th percentile) value, whereas all other uncertainties varied probabilistically. We did this by extracting the iterations with the relevant parameter values from the results for all iterations in the model run for the leading alternative. From this we calculated the expected value (i.e., mean) of the probability of persistence given low or high inputs for each parameter, respectively. The difference between the expected value for the low and the high input of each parameter is called the swing, with larger swings suggesting that the uncertainty in those parameters has more influence on the overall model results than those with smaller swings. These results were plotted as a tornado diagram (i.e., an inverted bar chart with the parameters ranked by their swing from highest to lowest). The parameters at the top of the tornado diagram (i.e., with the most swing) are those that the model is most sensitive to. Uncertainty in the parameters at the bottom of the tornado diagram have less influence on overall results. Although all uncertainties were included in the sensitivity analysis, we only plotted the top 10 uncertainties (i.e., those with the largest swings) in the tornado diagram.
We also used the tornado diagram to determine the “value of control” for each parameter (Johnson and Tani 2013). The value of control for each parameter identifies how valuable it would be if the uncertainty in that parameter could be resolved favorably. It is calculated as the difference between the overall expected value of the objective metric (i.e., probability of persistence) and the expected value if the parameter is held at its high or low value (whichever yields the preferable result). It can be read off the tornado diagram by subtracting the expected value from the highest value of the tornado bar for that parameter. For example, if the overall expected value is 0.4 and the expected value when a parameter is set at its high value is 0.7, then the value of control is 0.7 − 0.4 = 0.3.
Results
The chance of a successful reintroduction increased as the level of effort increased, as measured by the expected value of the probability of persistence (Figure 5). The Do Nothing alternative had no chance of success given that northern leopard frogs are currently extirpated from the region. For the remaining alternatives, the credible intervals were large, reflecting a wide range of epistemic uncertainty.
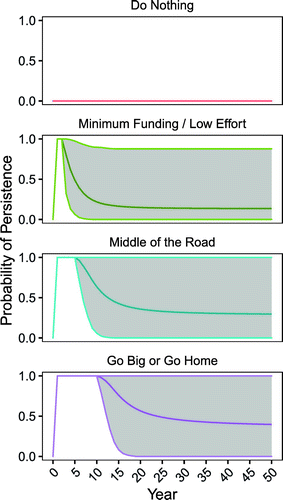
Probability of persistence estimates equaled 100% across all alternatives for the years in which releases occurred (Figure 5), reflecting the modelling assumption that at least some tadpoles will always survive to become young of year. The probability of persistence estimates decreased from this initial high value over time and stabilized approximately 10–20 years after the translocations ceased.
The Go Big or Go Home alternative had the largest expected value at year 50, with a mean predicted probability of persistence of 0.40 (Figure 6). Both the Go Big or Go Home and Middle of the Road alternatives were bimodal, as shown by the two wider areas at the ends of the violin plots in Figure 6, with many parameter combinations resulting in either very low (<5%) or relatively high (>95%) probabilities of a successful reintroduction and fewer parameter combinations with results in between.
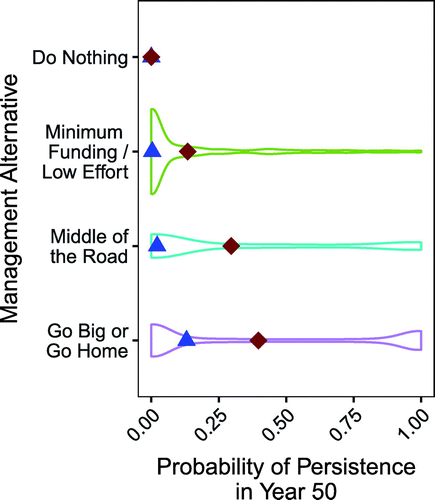
Note. Diamond, mean; triangle, median.
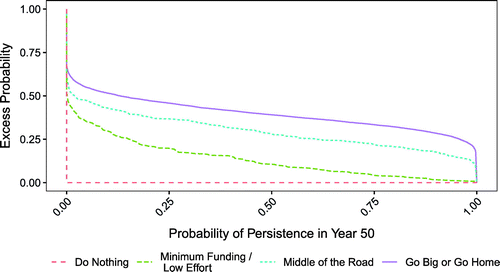
The Go Big or Go Home alternative was first-order stochastically dominant to the other alternatives (Figure 7; Appendix A). Specifically, the excess probabilities of the probability of persistence in year 50 for the Go Big or Go Home alternative were larger than those of the other alternatives for every value on the x axis (and therefore the lines of the excess probability distributions do not cross), indicating that it stochastically dominates all other alternatives. For example, consider the probability of persistence value of 0.25 on the x axis. The excess probability of Go Big or Go Home is approximately 0.5 (or 50%), indicating that there is a 50% chance that the probability of persistence in year 50 will be greater than 0.25. This is larger than the excess probabilities for that same x axis value for any of the other alternatives. The same holds true for every other value on the x axis.
The results of the sensitivity analysis using a tornado diagram for the leading alternative, Go Big or Go Home, are shown in Figure 8. The tornado diagram identifies the parameters in the model (recall the single ovals in Figure 3) for which epistemic uncertainty has the most influence on the results. The dashed vertical line at 0.4 in the tornado diagram shows the expected probability of persistence for the Go Big or Go Home alternative, which is the same as the mean value in Figure 6. The bars show the expected probability of persistence when each parameter is held at a high (≥90th percentile unless otherwise specified) or low (≤10th percentile unless otherwise specified) value, respectively, whereas all other parameters vary probabilistically. The parameter “egg survival rate reduction if bullfrog management is not effective” only applied in the model for runs where bullfrog management was not effective. Therefore, the reference value for this parameter is the value shown in the bar above for “bullfrog management effective–no” rather than the dotted black line. As a result, the high and low bars overlap and are both below the overall expected value of the probability of persistence.
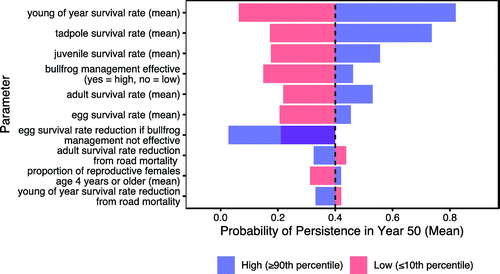
A key takeaway from the tornado diagram is that epistemic uncertainty in the survival estimates for all life stages ranked highly (i.e., had the largest swings), with mean survival for young of year and tadpoles being the two most influential epistemic uncertainties (Figure 8). These parameters also had the highest value of control, at approximately 0.42 and 0.34, respectively. The effectiveness of bullfrog management, as well as uncertainty in how bullfrogs affect egg survival, also ranked relatively highly on the tornado. Insights obtained from the tornado diagram led to further investigation of the top-ranking uncertainties (Randall et al. 2021).
Discussion
The results of our feasibility assessment suggest that a successful reintroduction in the BSCWMA is possible but far from certain. The leading alternative was Go Big or Go Home, with a mean probability of persistence of 0.4 at 50 years. Given that this alternative represents the highest level of effort, it is not surprising that it also has the highest probability of success. Although the probability of success is still relatively low for even this highest level of effort, it is consistent with previous amphibian reintroduction efforts; a review of published amphibian and reptile reintroductions found 41% had positive outcomes (Germano and Bishop 2009).
Using a decision-analytic approach to the modelling allowed us to calculate a range of potential outcomes around the mean probability of persistence and investigate how we could improve the probability of persistence. The distribution of the results for the leading alternative was bimodal, with the two modes being near the bounds of zero and one, suggesting that the reality is most likely one of two scenarios: either almost certain success or almost certain failure. Knowing which of these two scenarios is true would likely change the decision; for example, do it if success is almost certain, do not do it if failure is almost inevitable. Therefore, the decision maker may choose to gather more information before making a final decision. For example, a pilot study could focus on narrowing the uncertainty around the parameters at the top of the tornado diagram, that is, those that the model is most sensitive to: in this case, young of year and tadpole mean survival rates. Additionally, improved alternatives could be developed around parameters with the highest value of control, that is, those with uncertainty ranges with the greatest potential to increase success: in this case, also young of year and tadpole survival rates. We could brainstorm practical mitigation measures that would help control these key uncertainties: for example, head-starting tadpoles in captivity prior to release could improve tadpole survival and therefore improve the probability of a successful reintroduction.
Assessing stochastic dominance among alternatives is helpful for determining whether further work is needed to explore how the decision maker’s risk attitude could affect the decision (Howard 1966, Levy 1998, Johnson and Tani 2013; see Canessa et al. 2016 for a conservation example). If an alternative is stochastically dominant at the first order, as was the Go Big or Go Home alternative in this assessment, this alternative would be the rational choice regardless of the decision maker’s risk attitude and therefore no further work is needed to account for risk attitude in the analysis. If none of the alternatives showed first-order stochastic dominance (e.g., if one had a high mean probability of persistence but a large potential downside and another had a slightly lower mean probability of persistence but small potential downside), it would mean that risk attitude could affect the decision and be explored further (e.g., by assessing higher-order dominance (Canessa et al. 2016) or by explicitly assessing a utility function).
The decision maker will use the results of this feasibility assessment, in addition to other considerations such as cost, available funding, and current priorities, to inform their decision. Minimizing cost was identified as a fundamental objective, along with northern leopard frog recovery, during the framing steps of this assessment. The tradeoff between recovery and cost should be considered in a comprehensive analysis if they decide to move forward (Edwards et al. 2022). After further discussion with the decision maker, it became apparent that their preferred tradeoff between recovery and cost depended on the funding landscape; with more funding available (or less funding needed for other priorities), the decision maker would be more likely to proceed with the reintroduction at a lower probability of persistence. A measurable criterion that could be used in the future to capture this nuance between cost and the funding landscape is the proportion of the available budget, with lower proportions preferred.
Models are simplifications of reality: Their purpose is to provide useful insights, but insights should be interpreted in the context of model assumptions. In our model, we assumed that at least some tadpoles always survive to become young of year, reflecting the experience of our experts. If this is not true, then the probability of persistence would be lower because there would be more opportunities for the population to die out. We also assumed the same tadpole survival rates across all wetlands, regardless of the number of tadpoles in each wetland. Realistically, there would be density dependence in tadpole survival. We further assumed the same survival rate for all tadpoles, regardless of whether they were born in the wetlands or released as part of the translocation. Last, we did not use a quasi-extinction threshold in our model to calculate probability of persistence. Morris and Doak (2002) warn against using estimates of absolute probability of extinction because models may not account for factors such as inbreeding depression that result in unviable populations at low population sizes. Therefore, not using a quasi-extinction threshold may overestimate probability of persistence in our results. Further limitations and considerations are discussed in Randall et al. (2021).
The insights that come from models are also highly dependent on the quality of the model inputs. In this analysis, model inputs were parameterized using expert elicitation from a small number of in-house experts due to time constraints; this approach has been criticized in the past and resulted in recommendations for more comprehensive expert elicitation procedures (Keeney and von Winterfeldt 1991). Research has shown that including multiple experts with diverse perspectives, consulted through a formal elicitation protocol and with extensive training in probability judgements, can improve the quality of elicited information (Keeney and von Winterfeldt 1991, Hoffmann et al. 2007, Hemming et al. 2018, Hanea et al. 2021). Integrating these methods may have improved the quality of the model inputs.
We stress the importance of using modelling results for insights rather than absolute numbers (Johnson and Tani 2013). In general, population modeling results are useful for relative estimates (such as comparing alternatives) rather than as exact predictors of population viability (Morris and Doak 2002). The approach used in this feasibility assessment is intended to be used iteratively, with updates made to the quantitative forecasts as new information is obtained and new management alternatives are considered (e.g., adaptive management; Runge et al. 2020).
We found that this process was an effective way to integrate multiple perspectives from diverse stakeholders and resulted in a clear and transparent feasibility assessment that will be useful for decisions regarding northern leopard frog reintroduction. Although conservation practitioners may often worry about the difficulty of assessing cost, we describe here how it can still be useful to complete the recovery portion of a feasibility assessment while leaving the cost assessment for later. A lack of information regarding cost does not need to stop an analysis from proceeding; rather, the recovery portion of a feasibility assessment can be a useful first step from which the necessity for a cost analysis can be determined. Additionally, we found the sensitivity analysis around epistemic uncertainty to be particularly helpful for understanding which uncertainties are the most important. We emphasize that this approach can help conservation practitioners better understand their study system and direct future research efforts and/or conservation initiatives.
The overall process used here provides a decision-analytic framework useful for the evaluation of “whether” rather than simply “how” to undertake a conservation translocation. Future conservation translocations would benefit greatly from the widespread use of decision analysis to counter the complexity and uncertainty inherent in these decisions, providing a step-by-step process for what may otherwise be an overwhelming task. Even a relatively simple application of the process can provide insightful information (Gregory et al. 2012). Methods used here can be more broadly applied to conservation efforts for other species at a national or an international level to address the ongoing global biodiversity crisis.
The authors thank the participants who contributed to the problem framing, specification of objectives, and development of alternatives, including N. Allard, L. Anderson, M-A. Beaucher, S. Ehlers, C. Goldberg, L. A. Isaac, I. Manley, C. Peterson, and L. Svancara. The authors are grateful for feedback from Conor McGowan and two anonymous reviewers, whose comments and suggestions greatly improved the manuscript. The authors are also grateful for access to the computing cluster at the University of Idaho Research Computing and Data Services of the Institute for Interdisciplinary Data Sciences (RCDS of IIDS) to run the models. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Appendix A. Stochastic Dominance
When making decisions, decision makers respond to the available information according to their own risk attitude (or the risk attitude of the organization they represent). This means that different decision makers can respond differently to the same predictions, depending on whether they are risk-averse, risk-neutral or risk-seeking. Information about risk attitude is therefore important to assess rational strategies, but even in the absence of knowledge around the risk attitude of the decision makers, it is possible to gain insights into how important risk attitude will be to the decision using a method called stochastic dominance (Howard 1966, Levy 1998; see Canessa et al. 2016 for a conservation example).
Stochastic dominance can be determined by either comparing the cumulative probability distribution of each alternative or comparing the excess probability distribution (i.e., the complementary cumulative distribution). The cumulative probability distributions and the excess probability distributions convey the same information but require an inverse interpretation. The cumulative distribution plots the probability that a metric is less than or equal to the corresponding value on the x axis. The excess probability distribution plots the probability that a metric is greater than the corresponding value on the x axis. For objective metrics that we seek to maximize, alternatives with smaller cumulative probabilities (or larger excess probabilities) for a given value on the x axis are preferred as they represent a better chance of the objective metric being higher than the specified value. Alternatives with cumulative probabilities that are smaller than or equal to that of another alternative for every value on the x axis, which is the same as alternatives with excess probabilities that are larger than that of another alternative for every value on the x axis, are first-order stochastically dominant to the compared alternatives. Graphically, this first-order dominance is indicated by the fact that the lines of the probability distributions, cumulative or excess, do not cross (Figure 7). In other words, the dominant action would be the rational choice regardless of the decision maker’s risk attitude. Additional orders of dominance can be determined if needed, each requiring additional assumptions, but these extra steps were not required in our study.
Although the cumulative and excess probability distributions provide the same information, the excess probability distributions may be a better communication tool than cumulative distributions (Howard and Abbas 2015). Indeed, we found the excess probability distributions were more intuitive for our purposes. Therefore, we used excess probability distributions to plot our results.
References
- (2016) Alignment of threat, effort, and perceived success in North American conservation translocations. Conservation Biology 30(6):1159–1172.Crossref, Google Scholar
- (2021) Importance of species translocations under rapid climate change. Conservation Biology 35(3):775–783.Crossref, Google Scholar
- (2016) Stochastic dominance to account for uncertainty and risk in conservation decisions. Conservation Lett. 9:260–266.Crossref, Google Scholar
- (2014) Optimal release strategies for cost-effective reintroductions. J. Appl. Ecology 51(4):1107–1115.Crossref, Google Scholar
- (2018) Decision-making for mitigating wildlife diseases: From theory to practice for an emerging fungal pathogen of amphibians. J. Appl. Ecology 55(4):1987–1996.Crossref, Google Scholar
- (2013) Demographics of reintroduced populations: Estimation, modeling, and decision analysis. J. Wildlife Management 77(6):1081–1093.Crossref, Google Scholar
COSEWIC (2009) COSEWIC Assessment and Update Status Report on the Northern Leopard Frog Lithobates pipiens, Rocky Mountain Population, Western Boreal/Prairie Populations in Canada (Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ontario, Canada).Google Scholar- (2022) Trading off hatching success and cost in the captive breeding of Whooping Cranes. Animal Conservation 25(1):101–109.Crossref, Google Scholar
- (2010) Population viability analysis: Origins and contributions. Nature Ed. Knowledge 3(10):15.Google Scholar
- (2009) Distribution and postbreeding environmental relationships of northern leopard frogs (Rana [Lithobates] Pipiens) in Washington. Western North Amer. Nature 69(4):537–547.Crossref, Google Scholar
- (2009) Suitability of amphibians and reptiles for translocation. Conservation Biology 23(1):7–15.Crossref, Google Scholar
- (2012) Structured Decision Making: A Practical Guide to Environmental Management Choices (Wiley-Blackwell, Chichester, West Sussex, United Kingdom).Crossref, Google Scholar
- (1999) Smart Choices: A Practical Guide to Making Better Decisions (Harvard Business School Press, Boston).Google Scholar
- (2021) Uncertainty quantification with experts: Present status and research needs. Risk Anal. 42(2):254–263.Google Scholar
- (2016) Developments in amphibian captive breeding and reintroduction programs. Conservation Biology 30(2):340–349.Crossref, Google Scholar
- (2020) Weighting and aggregating expert ecological judgments. Ecology Appl. 30(4):e02075.Crossref, Google Scholar
- (2018) Eliciting improved quantitative judgements using the IDEA protocol: A case study in natural resource management. PLoS One. 13(6):e0198468.Crossref, Google Scholar
- (2022) An introduction to decision science for conservation. Conservation Biol. 36(1):1–16.Crossref, Google Scholar
- (2019) Predicting reintroduction costs for wildlife populations under anthropogenic stress. J. Appl. Ecology 57(1):192–201.Google Scholar
- (1981) Leopard Frog Populations and Mortality in Wisconsin, vol. 122 (Wisconsin Department of Natural Resources, Madison, WI).Google Scholar
- (2004) Evolutionary history of the northern leopard frog: reconstruction of phylogeny, phylogeography, and historical changes in population demography from mitochondrial DNA. Evolution 58(1):145–159.Google Scholar
- (2007) Elicitation from large, heterogeneous expert panels: Using multiple uncertainty measures to characterize information quality for decision analysis. Decision Anal. 4(2):91–109.Link, Google Scholar
- (1966) Decision Analysis: Applied Decision Theory (John Wiley, New York).Google Scholar
- (2015) Foundations of Decision Analysis (Pearson Education, Boston).Google Scholar
- (2005) Influence diagrams. Decision Anal. 2(3):127–143.Link, Google Scholar
Idaho Department of Fish and Game (2017) Idaho State Wildlife Action Plan, 2015 (Idaho Department of Fish and Game, Boise).Google ScholarIUCN/SSC (2013) Guidelines for reintroductions and other conservation translocations. Accessed August 1, 2021, http://www.issg.org/pdf/publications/translocation-Guidelines-2012.pdfGoogle Scholar- (2013)
Perform probabilistic analysis and identify insights . Parnell GS, Tani SN, Johnson ER, eds. Handbook of Decision Analysis (John Wiley & Sons, Hoboken, NJ), 248–290.Crossref, Google Scholar - (2016) The metalog distributions. Decision Anal. 13(4):243–277.Link, Google Scholar
- (1991) Eliciting probabilities from experts in complex technical problems. IEEE Trans. Engrg. Management 38(3):191–201.Crossref, Google Scholar
- (2017) A decision-theory approach to cost-effective population supplementation for imperiled species. Ecology Econom. 142:194–202.Crossref, Google Scholar
- (1998) Stochastic Dominance: Investment Decision Making Under Uncertainty (Kluwer Academic Publishers, The Netherlands).Crossref, Google Scholar
- (2016) Multi-species baseline initiative. Project report. Idaho Department of Fish and Game, Coeur d’Alene, ID. https://idfg.idaho.gov/species/bibliography/1510084.Google Scholar
- (2020) Status of pond-breeding amphibians in Northern Idaho and Northeastern Washington, USA. Herpetology Conservation Biology 15(3):526–535.Google Scholar
- (2018) Prioritizing recovery funding to maximize conservation of endangered species. Conservation Lett. 11(6):e12604.Crossref, Google Scholar
- (2011) Incorporating parametric uncertainty into population viability analysis models. Biology Conservation 144(5):1400–1408.Crossref, Google Scholar
- (2007) Decision Analysis for the Professional (SmartOrg, Inc., Menlo Park, CA).Google Scholar
- (1968) A comparison of the estimated size and the “effective size” of breeding populations of the Leopard Frog, Rana pipiens. Evolution 22(2):274–283.Google Scholar
- (2002) Quantitative Conservation Biology (Sinauer Associates, Sunderland, MA).Google Scholar
NatureServe (2022) NatureServe Explorer. Accessed April 20, 2022, https://explorer.natureserve.org/.Google ScholarR Core Team (2021) R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, Vienna, Austria).Google Scholar- (2021) Feasibility Assessment for Recovery of the Northern Leopard Frog (Lithobates pipiens) in the Kootenai Valley, Idaho (Wilder Institute/Calgary Zoo, Calgary, Alberta, Canada).Google Scholar
- (2005)
Rana pipiens Schreber, 1782. Northern leopard frog . Lannoo MJ, ed. Amphibian Declines: The Conservation Status of United States Species (University of California Press, Berkeley, CA), 570–577.Google Scholar - (2020) Structured Decision Making: Case Studies in Natural Resource Management (Johns Hopkins University Press, Baltimore).Crossref, Google Scholar
- (2020) Integrating genetics and metapopulation viability analysis to inform translocation efforts for the last northern leopard frog population in Washington State, USA. J. Herpetology 54(4):465–475.Crossref, Google Scholar
- (2007) Developing the science of reintroduction biology. Conservation Biology 21:303–312.Crossref, Google Scholar
- (2021) Global Conservation Translocation Perspectives: 2021. Case Studies from Around the Globe (IUCN SSC Conservation Translocation Specialist Group, Environment Agency, Abu Dhabi & Calgary Zoo, Calgary, Alberta, Canada).Google Scholar
- (2010) Reducing overconfidence in the interval judgments of experts. Risk Anal. 30(3):512–523.Crossref, Google Scholar
- (2018) Projecting further increases in conservation translocations: A Canadian case study. Biology Conservation 228:175–182.Crossref, Google Scholar
- (2013a)
Design creative alternatives . Parnell GS, Tani SN, Johnson ER, eds. Handbook of Decision Analysis (John Wiley & Sons, Hoboken, NJ), 149–165.Crossref, Google Scholar - (2013b)
Frame the decision opportunity . Parnell GS, Tani SN, Johnson ER, eds. Handbook of Decision Analysis (John Wiley & Sons, Hoboken, NJ), 110–126.Crossref, Google Scholar - (2014) Boundary-Smith Creek Wildlife Management Area: 2014–2023 Management Plan. (Idaho Department of Fish and Game, Coeur d’Alene, ID). https://idfg.idaho.gov/sites/default/files/2014-2023-BoundarySCWMA-Plan-Final.pdf.Google Scholar
- (1997) Structured-Population Models in Marine, Terrestrial, and Freshwater Systems (Chapman and Hall, New York).Crossref, Google Scholar
- (2000)
Population viability analysis: Data requirements and essential analyses . Boitani L, Fuller TK, eds. Research Techniques in Animal Ecology: Controversies and Consequences (Columbia University Press, New York), 288–331.Google Scholar
Laura M. Keating is a conservation decision analyst at the Wilder Institute/Calgary Zoo. She holds a BScH in environmental science/biology from Queen’s University and an MSc in mathematical modelling and scientific computing from the University of Oxford. She previously worked in both environmental and corporate strategy consulting, the latter at Strategic Decisions Group where she received training in decision analysis. She is a certified practitioner with the Society of Decision Professionals.
Lea Randall is the interim senior manager of conservation operations and has worked as a population ecologist at the Wilder Institute/Calgary Zoo for the last 11 years. She earned her BSc in biology from the University of Victoria in 2005 and her MSc in ecology from the University of Calgary in 2009. Her primary research focuses on the population dynamics, genetics, reproduction, and reintroduction of Northern Leopard Frogs.
Rebecca Stanton is a conservation research associate at the Wilder Institute/Calgary Zoo. She completed her BSc in Biology at Wilkes University and her MSc in biology at the University of Saskatchewan. Her primary research focuses on reintroduction of northern leopard frogs.
Casey McCormack is a regional wildlife diversity biologist for Idaho Department of Fish and Game in the Idaho Panhandle. He earned his BS degree in wildlife resources and MS degree in environmental science from the University of Idaho. His work and research interests include habitat preservation, habitat restoration, and climate adaptation for sensitive species in western North America.
Michael Lucid is a biologist and co-owner of Selkirk Wildlife Science, LLC, Sandpoint, ID. He was a wildlife diversity biologist with the Idaho Department of Fish and Game from 2009 to 2020. He is a 2017 Wilburforce fellow in conservation science and received an MS degree in biology from Idaho State University, Pocatello, ID.
Travis Seaborn is an assistant professor at North Dakota State University in the School of Natural Resource Sciences. He earned his PhD from the School of Biological Sciences at Washington State University. His research program is focused on the conservation and management of species impacted by humans within various social-ecological systems. He uses genomics, landscape ecology, and a variety of modeling approaches, including individual-based models, to investigate these systems.
Sarah J. Converse is the unit leader of the U.S. Geological Survey, Washington Cooperative Fish and Wildlife Research Unit, and an associate professor at the University of Washington in Seattle, WA. Her research focuses on the development and application of methods to understand the functioning of populations, especially small and declining populations, and to improve their management.
Stefano Canessa is a senior postdoctoral fellow in the Division of Conservation Biology at the University of Bern, Switzerland. He earned a PhD in conservation biology from the University of Melbourne in 2016. His research focuses on demographic modeling, risk analysis and decision making for endangered species management, especially conservation translocations. He has been involved in species recovery plans across the globe, ranging from frogs and turtles to birds and bats.
Axel Moehrenschlager serves on the Leadership and Steering Committee of International Union for Conservation of Nature’s (IUCN) Species Survival Commission (SSC) and is the chair of the IUCN SSC Conservation Translocation Specialist Group. He is an adjunct professor at the University of Calgary in Canada, adjunct associate professor at Clemson University in the United States, Erskine fellow at the University of Canterbury in New Zealand, and research associate at Oxford University, where he received his DPhil.